Publié le 22 mar 2009Lecture 8 min
Physiopathologie de la migraine en 2009
Christian LUCAS, Clinique neurologique, CHRU de Lille
La compréhension de la physiopathologie de la migraine a beaucoup évolué au cours de ces dernières décennies. Rappelons que dans la théorie vasculaire développée par H.G.Wolff (1), l’aura migraineuse était expliquée par une vasoconstriction primitive transitoire, et la céphalée par une vasodilatation de rebond des artères méningées provoquant l’activation des nocicepteurs périvasculaires1.Puis,dans la théorie neuronale,l’aura est expliquée par la survenue d’une dépression corticale envahissante (spreading depression de A.P.Leao (2)), avec implication ensuite du circuit trigéminovasculaire et libération de différents neuropeptides pro-inflammatoires,dont le CGRP (Calcitonin Gene Related Peptide),le tout étant sous-tendu par une susceptibilité génétique. Les modèles cliniques quasi-expérimentaux de la migraine hémiplégique familiale,affection autosomique dominante, font désormais considérer la migraine comme une possible maladie des canaux ioniques («channelopathy ») permettant ainsi d’expliquer l’aspect paroxystique,évoluant par crise de la maladie.
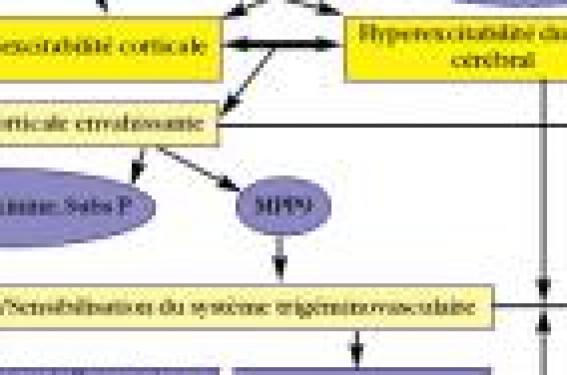
La DCE ou « spreading depression » La dépression corticale envahissante (DCE) correspond à une vague de dépolarisation neuronale et gliale suivie d’une dépression neuronale de longue durée, observée expérimentalement après application de potassium sur les cortex de rats. Ce phénomène électrique est contemporain d’une vague d’hypoperfusion,puis quelques heures plus tard,d’une hyperperfusion cérébrale. Cette DCE progresse lentement à la surface du cortex et peut être interrompue par un sillon ou une scissure. La DCE induit une hyperactivité neuronale initiale avec libération massive de K+ et de glutamate,suivie d’une entrée massive de Na+ et de Ca++ dans les neurones et les astrocytes.Les astrocytes, en situation d’hypofonctionnement, sont alors incapables de protéger le neurone du glutamate et du K+ qui sont en excès dans le milieu extracellulaire, avec pérennisation de la dépolarisation neuronale. Le glutamate joue un rôle essentiel avec le blocage de la DCE par les antagonistes des récepteurs NMDA. Chez l’homme,le rôle de la DCE est resté longtemps controversé du fait de la rareté de déclenchement des DCE et des difficultés à interpréter les phénomènes électromagnétiques observés. Grâce à l’utilisation de stimulations visuelles répétées prolongées en IRMf par effet bold, Y.Cao et coll. (3) ont pu démontrer que certaines zones cérébrales passent d’un état de réponse à la stimulation visuelle vers un état de non-réponse selon un « front de désactivation »,se propageant avec le temps le long de la scissure calcarine à une vitesse de 3 à 6 mm/min.Ces données ont été confirmées lors de l’étude d’auras migraineuses spontanées par l’équipe de N.Hadjikhani (4). Ces phénomènes ne semblent toutefois pas spécifiques à l’aura, puisqu’ils peuvent parfois s’observer au cours de crises migraineuses sans aura (5). La DCE semble donc bien être à l’origine de l’aura migraineuse, et est vraisemblablement aussi impliquée dans la migraine sans aura, soit avec une DCE survenant dans des zones silencieuses, soit avec une perturbation de l’excitabilité corticale mais sans traduction clinique. Cela pose la question d’une seule et même maladie migraineuse, sans et avec aura, et la question d’une physiopathologie monoforme ou non. Il existe de façon clairement démontrée une hyperexcitabilité neuronale (6) dans la migraine,notamment du cortex occipital avec une pauvreté de la neurotransmission inhibitrice (faible densité en récepteurs GABA), une richesse de la neurotransmission excitatrice (densité très élevée en récepteurs NMDA) et une synchronisation fonctionnelle élevée (densité neuronale élevée et haut niveau d’architecture fonctionnelle,densité élevée en jonctions GAP, faible densité relative en astrocytes). Les migraineux ont également, lors d’enregistrements de potentiels évoqués multimodaux, un défaut d’habituation,c'est-à-dire la diminution de l’amplitude des potentiels normalement observés lorsque l’on répète les trains de stimulations. Différents noyaux du tronc cérébral (5,7) sont impliqués comme générateurs ou modulateurs dans la crise de migraine.Récemment,l’équipe française de G.Géraud (8) a montré,après soulagement total de la céphalée migraineuse,une persistance en TEP et en IRM d’une activation dans le tronc cérébral (mésencéphale et partie haute du bulbe) et de l’hypothalamus, suggérant que ces structures pourraient être des génératrices de la crise. La persistance de cette activation pourrait peut-être expliquer les phénomènes de récurrences des crises.Il pourrait s’agir de zones clés situées à l’interface entre les systèmes excitateurs et les systèmes inhibiteurs de la nociception céphalique. Une hyperexcitabilité des neurones thalamo-corticaux et cortico-corticaux est présente en période de crise (expliquant la phono-,photophobie,l’osmophobie, l’allodynie tactile et thermique). Nous avons fait état plus haut des phénomènes d’hyperexcitabilité du cortex,notamment occipital. La primauté du tronc cérébral ou du cortex comme « générateur » de la crise migraineuse n’est pas établie à ce jour, sachant que les deux mécanismes sont loin d’être antithétiques et que ces structures pourraient avoir un rôle permissif sur le déclenchement de la crise sans être le(s) générateur(s) princeps. Système trigémino-vasculaire et inflammation neurogène Selon la théorie neuro-trigéminovasculaire de M.A.Moschowitz (9), la stimulation de nocicepteurs périvasculaires de la dure-mère et des méninges et/ou les nocicepteurs cervicaux active les neurones du noyau trigéminal caudal.Les influx nerveux générés seraient relayés de façon orthodromique jusqu’au thalamus puis au cortex,et permettraient l’élaboration de la perception consciente de douleur, tandis que l’activation antidromique des fibres trigéminales amyéliniques entraîne une libération de neuropeptides (SP,CGRP,neurokinine A). Les relations entre la DCE, l’aura et la céphalée migraineuse restent incertaines. Des travaux expérimentaux (9) sont en faveur du déclenchement des céphalées par la DCE survenant en phase d’aura. Ils montrent que la répétition de la DCE induit une augmentation de l’activité neuronale dans le noyau caudal du trijumeau,élément clé dans la genèse de la sensation douloureuse. Données récentes sur DCE, gènes, hormones et canaux ioniques Des données remarquables sur DCE, gènes,hormones et canaux ioniques viennent juste d’être publiées par l’équipe de M.A Moschkowitz (10),avec des rappels sur les dysfonctionnements des canaux calciques et des pompes en rapport avec des mutations génétiques.La traduction clinique de ce dysfonctionnement ionique est illustrée par la migraine hémiplégique familiale (MHF),dont trois types sont à l'heure actuelle connus.Cette maladie représente un lien entre l'implication des gènes et la DCE. Au cours de cette pathologie,les crises sont déclenchées par le stress,l'effort,un traumatisme même minime. Les crises se manifestent par des migraines avec aura et le nombre de crises serait plus important chez la femme. La MHF est en rapport avec une mutation sur le gène CACNA1A pour le type 1 et avec une mutation sur la sous-unité α 2 de la pompe ATP-ase dépendante pour le type 2. Les variations de susceptibilité de la DCE entre les deux sexes ont été testées chez un modèle de souris (R122QK1), avec l'hypothèse que l’imprégnation hormonale pouvait expliquer une proportion plus importante de crises chez la femme. Physiopathologie présumée de la migraine. Il a été démontré que l'ovariectomie entraînait chez la souris une diminution de la survenue de la DCE. Les mêmes résultats ont été reproduits chez la souris ménopausée.Il existe donc chez l'animal des preuves d'une relation entre les hormones ovariennes et la DCE. Les traitements de fond de la migraine pourraient agir en supprimant la DCE. Dans le travail rapporté par C.Ayata en 200611, cinq substances ont été essayées chez l'animal (valproate,topiramate, bêtabloquants,amitriptylline et méthylgergide),prouvant avec un délai de plusieurs semaines que ces substances diminuent la DCE. Enfin,les travaux de T.Takano (12) sur la NADH (nicotinamide adénine dinucléotide), substance responsable du transfert d'énergie dans la chaîne mitochondriale et mesurée par techniques fluorométriques,ont démontré que l'hypoxie était un facteur clé du déclenchement de la DCE. La DCE induirait une cascade biochimique conduisant à la rupture de la barrière hémato-encéphalique par un mécanisme activateur dépendant de la métalloprotéinase MMP9. Ces données suggèrent de possibles risques lésionnels liés à la survenue d’une aura migraineuse (hypersignaux de substance blanche ?). Elles ouvrent également la voie à de nouvelles pistes thérapeutiques :certains inhibiteurs de l’activité métalloprotéinase pourraient être utilisés. Ces travaux remarquables de l’équipe de M.A.Mosckowitz permettent de mieux articuler les différents éléments du puzzle de la physiopathologie de la migraine (figure).
Attention, pour des raisons réglementaires ce site est réservé aux professionnels de santé.
pour voir la suite, inscrivez-vous gratuitement.
Si vous êtes déjà inscrit,
connectez vous :
Si vous n'êtes pas encore inscrit au site,
inscrivez-vous gratuitement :




